
O glucagon é um hormônio peptídico, produzido pelas células alfa do pâncreas. O glucagon aumenta a concentração de glicose e ácidos graxos na corrente sanguínea, sendo considerado o principal hormônio catabólico do corpo. O glucagon também é utilizado como medicação para tratar uma série de condições de saúde. O efeito do glucagon é oposto ao da insulina, que reduz a glicose extracelular. O glucagon é produzido a partir do proglucagon, codificado pelo gene GCG.
O pâncreas libera glucagon quando a quantidade de glicose na corrente sanguínea está muito baixa. O glucagon faz com que o fígado entre em glicogenólise: convertendo o glicogênio armazenado em glicose, que é liberada na corrente sanguínea. Níveis elevados de glicose no sangue, por outro lado, estimulam a liberação de insulina. A insulina permite que a glicose seja absorvida e utilizada por tecidos que dependem de insulina. Assim, o glucagon e a insulina fazem parte de um sistema de feedback que mantém os níveis de glicose no sangue estáveis. O glucagon aumenta o gasto de energia e está elevado em condições de estresse. O glucagon pertence à família de hormônios secretina.
Função do glucagon
O glucagon geralmente eleva a concentração de glicose no sangue promovendo a gluconeogênese e a glicogenólise. O glucagon também diminui a síntese de ácidos graxos nos tecidos adiposos e no fígado. O glucagon promove a lipólise nesses tecidos, o que faz com que liberem ácidos graxos na circulação, onde podem ser catabolizados para gerar energia em tecidos como o músculo esquelético, quando necessário.
A glicose é armazenada no fígado na forma do polissacarídeo glicogênio, que é um glucano (um polímero composto por moléculas de glicose). As células do fígado (hepatócitos) possuem receptores de glucagon. Quando o glucagon se liga aos receptores de glucagon, as células do fígado convertem o glicogênio em moléculas individuais de glicose e as liberam na corrente sanguínea, em um processo conhecido como glicogenólise. Quando o armazenamento de glicose se esgota, o glucagon incentiva o fígado e os rins a sintetizar glicose adicional pela gluconeogênese. O glucagon desativa a glicólise no fígado, fazendo com que intermediários glicolíticos sejam direcionados para a gluconeogênese.
O glucagon também regula a taxa de produção de glicose através da lipólise. O glucagon induz a lipólise em humanos sob condições de supressão da insulina (como no diabetes mellitus tipo 1).
A produção de glucagon parece depender do sistema nervoso central através de vias ainda não definidas. Em animais invertebrados, a remoção do pedúnculo ocular foi relatada como afetando a produção de glucagon. A excisão do pedúnculo ocular em jovens camarões de água doce provoca hiperglicemia induzida por glucagon.
Mecanismo de ação do glucagon

O glucagon se liga ao receptor de glucagon, um receptor acoplado a proteína G, localizado na membrana plasmática da célula. A mudança de conformação no receptor ativa as proteínas G, uma proteína heterotrimérica com subunidades α, β e γ. Quando a proteína G interage com o receptor, ela sofre uma mudança conformacional que resulta na substituição da molécula de GDP que estava ligada à subunidade α por uma molécula de GTP. Essa substituição resulta na liberação da subunidade α das subunidades β e γ. A subunidade alfa ativa especificamente a próxima enzima na cascata, a adenilato ciclase.
A adenilato ciclase fabrica adenosina monofosfato cíclico (AMP cíclico ou cAMP), que ativa a proteína quinase A (quinase dependente de cAMP). Essa enzima, por sua vez, ativa a fosforilase quinase, que depois fosforila a fosforilase de glicogênio b (PYG b), convertendo-a na forma ativa chamada fosforilase a (PYG a). A fosforilase a é a enzima responsável pela liberação de glicose 1-fosfato dos polímeros de glicogênio.
Um exemplo da via seria quando o glucagon se liga a uma proteína transmembrana. As proteínas transmembrana interagem com Gɑβ𝛾. Gɑ se separa de Gβ𝛾 e interage com a proteína transmembrana adenilato ciclase. A adenilato ciclase catalisa a conversão de ATP em cAMP. O cAMP se liga à proteína quinase A, e o complexo fosforila a fosforilase quinase. A fosforilase quinase fosforilada fosforila a fosforilase. A fosforilase fosforilada clipe unidades de glicose do glicogênio como glicose 1-fosfato.
Adicionalmente, o controle coordenado da glicólise e da gluconeogênese no fígado é ajustado pelo estado de fosforilação das enzimas que catalisam a formação de um potente ativador da glicólise chamado frutose 2,6-bisfosfato. A enzima proteína quinase A (PKA), que foi estimulada pela cascata iniciada pelo glucagon, também fosforila um único resíduo de serina da cadeia polipeptídica bifuncional contendo tanto as enzimas frutose 2,6-bisfosfatase quanto a fosfofrutoquinase-2. Essa fosforilação covalente iniciada pelo glucagon ativa a primeira e inibe a segunda. Isso regula a reação que catalisa o frutose 2,6-bisfosfato (um potente ativador da fosfofrutoquinase-1, a enzima que é o principal passo regulatório da glicólise) reduzindo a taxa de sua formação, inibindo assim o fluxo pela via da glicólise e permitindo que a gluconeogênese predomine. Esse processo é reversível na ausência de glucagon (e portanto, na presença de insulina).
A estimulação do glucagon pela PKA também inativa a enzima glicolítica piruvato quinase nos hepatócitos.
Fisiologia
Produção de glucagon
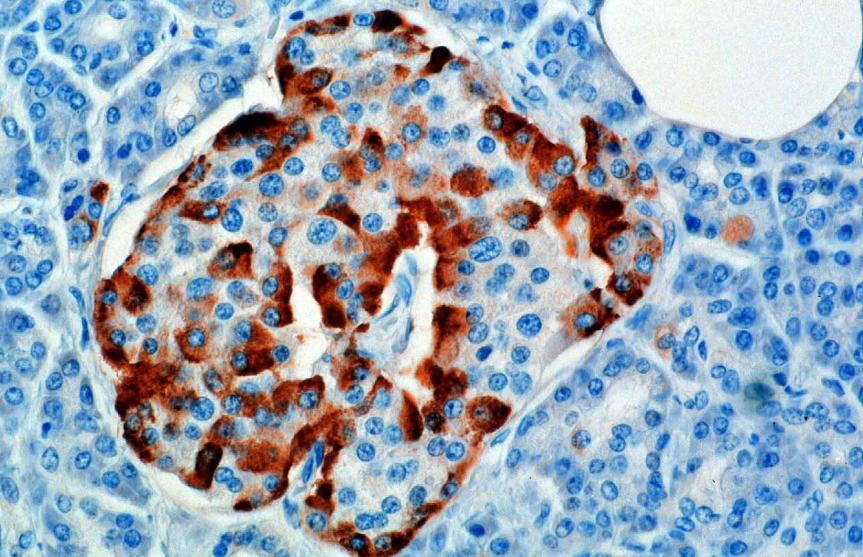
O hormônio é sintetizado e secretado pelas células alfa (células α) das ilhotas de Langerhans, que estão localizadas na parte endócrina do pâncreas. A produção, que por outro lado é livremente contínua, é suprimida/regulada pela amiloide, um hormônio peptídico co-secretado com insulina pelas células β do pâncreas. À medida que os níveis de glicose no plasma diminuem, a redução subsequente na secreção de amiloide alivia sua supressão das células α, permitindo assim a secreção de glucagon.
Em roedores, as células alfa estão localizadas na borda externa da ilhota. A estrutura da ilhota humana é muito menos segregada, e as células alfa estão distribuídas por todo o ilhotas em estreita proximidade com as células beta. O glucagon também é produzido por células alfa no estômago.
Pesquisas recentes demonstraram que a produção de glucagon também pode ocorrer fora do pâncreas, sendo o intestino o local mais provável de síntese extrapancreática de glucagon.
Regulação do glucagon
A secreção de glucagon é estimulada por:
- Hipoglicemia
- Epinefrina (através dos receptores adrenérgicos β2, α2 e α1)
- Arginina
- Alanina (frequentemente derivada da transaminação do piruvato/glutamato muscular)
- Acetilcolina
- Colescistoquinina
- Peptídeo inibidor gástrico
A secreção de glucagon é inibida por:
- Somatostatina
- Amiloide
- Insulina (através do GABA)
- PPARγ/receptor X retinoide heterodímero.
- Aumento de ácidos graxos livres e ácidos cetônicos no sangue.
- Aumento da produção de ureia
- Peptídeo semelhante ao glucagon-1
Estrutura do glucagon
O glucagon é um polipeptídeo de 29 aminoácidos. Sua estrutura primária em humanos é: NH2-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp-Leu-Met-Asn-Thr-COOH.
O polipeptídeo tem uma massa molecular de 3485 daltons. O glucagon é um hormônio peptídico (não esteroide).
O glucagon é gerado a partir do clivagem do proglucagon pela proproteína convertase 2 nas células alfa das ilhotas pancreáticas. Nas células L intestinais, o proglucagon é clivado em produtos alternativos como a glicentina, GLP-1 (um incretina), IP-2, e GLP-2 (que promove o crescimento intestinal).
Patologia
Níveis anormalmente elevados de glucagon podem ser causados por tumores pancreáticos, como o glucagonoma, cujos sintomas incluem eritema migratório necrólitico, redução de aminoácidos e hiperglicemia. Pode ocorrer isoladamente ou no contexto da neoplasia endócrina múltipla tipo 1.
O glucagon elevado é o principal responsável pela cetoacidose hiperglicêmica em diabetes tipo 1 não diagnosticado ou mal tratado. À medida que as células beta param de funcionar, a insulina e o GABA pancreático não estão mais presentes para suprimir a produção livre de glucagon. Como resultado, o glucagon é liberado das células alfa em sua máxima capacidade, provocando uma rápida quebra do glicogênio em glicose e uma rápida cetogênese. Foi constatado que um subconjunto de adultos com diabetes tipo 1 leva 4 vezes mais tempo, em média, para desenvolver cetoacidose quando recebem somatostatina (inibe a produção de glucagon) sem insulina. A inibição do glucagon tem sido uma ideia popular para o tratamento do diabetes, no entanto, alguns alertaram que fazê-lo pode dar origem a diabetes instável em pacientes com glicose no sangue adequadamente estável.
A ausência de células alfa (e, portanto, de glucagon) é considerada uma das principais influências na extrema volatilidade da glicose no sangue no contexto de uma pancreatectomia total.
História
Na década de 1920, Kimball e Murlin estudaram extratos pancreáticos e encontraram uma substância adicional com propriedades hiperglicêmicas. Eles descreveram o glucagon em 1923. A sequência de aminoácidos do glucagon foi descrita no final da década de 1950. Um entendimento mais completo de seu papel na fisiologia e na doença não foi estabelecido até a década de 1970, quando foi desenvolvido um radioimunoensaio específico.